Species :
1- Oophaga pumilio (Schmidt, 1857) — Strawberry poison-dart frog







From Wikipedia, the free encyclopedia
The strawberry poison frog or strawberry poison-dart frog (Oophaga pumilio, formerly Dendrobates pumilio) is a species of small poison dart frog found in Central America.[2] It is common throughout its range, which extends from eastern central Nicaragua through Costa Ricaand northwestern Panama. The species is often found in humid lowlands and premontane forest, but large populations are also found in disturbed areas such as plantations.[3] The strawberry poison frog is perhaps most famous for its widespread variation in coloration, comprising approximately 15–30 color morphs, most of which are presumed to be true-breeding.[4] O. pumilio, while not the most poisonous of the dendrobatids, is the most toxic member of its genus.
Strawberry poison-dart frog





Conservation status :
Least Concern (IUCN 3.1)[1]
Scientific classification:
Kingdom:Animalia
Phylum:Chordata
Class:Amphibia
Order:Anura
Family:Dendrobatidae
Genus:Oophaga
Species:O. pumilio
Binomial name:
Oophaga pumilio
(Schmidt, 1857)
Synonyms :
Dendrobates pumilio Schmidt, 1857
Diet :
The diet of O. pumilio causes the skin of the amphibian to become toxic in nature when certain subspecies of mites and ants are ingested.[5][6] Alkaloid toxins are organic in nature and contain nitrogenous bases that react with carbon and hydrogen groups.[7] Pumiliotoxin 251D is the specialized toxin that is sequestered by this species of frog. This toxin has a negative stimulating effect on cardiac function and is a severe disruptor of the sodium potassium ion channels within cells. Upon ingestion of Pumiliotoxin 251D, organisms preying on O. pumilioexperience convulsions, paralysis, and death.[7]
Oribatida mites belonging to the glandulate suborder Brachypylina are an important origin of pumiliotoxins in O. pumilio. Hexane-extraction techniques indicate presence of alkaloid toxins in Brachypylina.[8] Toxins appear to be biosynthesized in adult mites, as nymph and larval stages of the arachnid do not carry the toxins. Experimental analysis of this species of mite show alkaloid toxins are found almost exclusively in the opisthonotal glands of mites of the Scheloribatidae.[9] Oil glands of the mite contain the toxins and are then released internally as the amphibian digests the arthropod.
O. pumilio can also attribute its cutaneous toxicity to its rich diet of formicine ants.[6] Species of the formicine genus Brachymyrmex contain pumiliotoxins which the frogs incorporate and accumulate poison from.[10] There is a variability of alkaloid profiles among populations and individuals of O. pumilio, which is indicative of varying levels of available prey within their intraspecific habitats.[11] Research and physical analysis reveal that maternally derived alkaloids exist in young tadpoles.[12] During tadpole-rearing, mother frogs feed their young an unfertilized egg from their ovaries after dropping each individual tadpole into a repository of water usually found in a bromeliad.[13] Tadpoles lacking the obligate nutritive egg diet do not contain the alkaloid.[12] This step is crucial for the tadpoles to sequester the alkaloid from their mother; without such, young tadpoles become susceptible to predation by arthropods and other frogs.
Behavior :
Oophaga pumilio is diurnal and primarily terrestrial, and can often be found in leaf litter in both forested and disturbed areas. Though brightly colored and toxic, these frogs are relatively small, growing to approximately 17.5–22 mm (0.69–0.87 in) in standard length.[3] Males are extremely territorial, guarding small territories;[14] females and juveniles are far more sociable.
Reproduction and parental care :
Oophaga pumilio is an external breeder, and other species of the genus Oophaga are notable in the amphibian world for exhibiting a high degree of parental care.[15] The strawberry poison frog has dual parental care.[16] The males defend and water the nests, and the females feed the oophagous tadpoles their unfertilized eggs. Although both male and female contribute to parental care, females invest more heavily in terms of energy expenditure, time investment, and loss of potential reproduction.[16] Females provide energetically costly eggs to the tadpoles for 6–8 weeks (until metamorphosis), remain sexually inactive during tadpole rearing, and care for only one clutch of four to 6 tadpoles at a time.[16] The males, though, contribute the relatively "cheap" (in terms of energy) act of watering and protecting the eggs for a relatively short period (10–12 days), and can care for multiple nests at one time.[16]The extreme maternal investment in their offspring is believed to be the result of high egg mortality. Only 5–12% of the clutch develops into tadpoles, so the female's fitness may be best increased by making sure those few eggs that form tadpoles survive.[17]
After mating, the female lays three to five eggs on a leaf or bromeliad axil. The male then ensures the eggs are kept hydrated by transporting water in his cloaca. After about 10 days, the eggs hatch and the female transports the tadpoles on her back to some water-filled location.[18]In captivity, on rare occasions, the male is observed transporting the tadpoles, though whether this is intentional, or the tadpoles simply hitch a ride, is unknown. Bromeliad axils are frequently used tadpole deposition sites, but anything suitable can be used, such as knots in trees, small puddles, or human trash such as aluminum cans.
Tadpoles are deposited singly at each location. Once this has been done, the female will come to each tadpole every few days and deposit several unfertilized food eggs.[3] In captivity, tadpoles have been raised on a variety of diets, ranging from algae to the eggs of other dart frogs, but with minimal success. O. pumilio tadpoles are considered obligate egg feeders, as they are unable to accept any other form of nutrition.
After about a month, the tadpole will metamorphose into a small froglet. Generally, it stays near its water source for a few days for protection as it absorbs the rest of its tail.
The la gruta morph from Colón Province, Panama
Taxonomy :
Oophaga pumilio belongs to the genus Oophaga,[15] although the name Dendrobates pumilio is still sometimes used. There is evidence that the species of the Oophaga genus (previously classified as the "female parental care group" of Dendrobates[19]) are a monophyletic evolutionary group. Due to the low level of genetic divergence between the species analyzed in this genus, it is estimated that they speciated relatively recently, after the formation of the current Panamanian land bridge in the Pliocene (3–5 million years ago).[20]Oophaga pumilio is believed to be most closely related to Oophaga arborea.[21]
Evolution :
Strawberry poison frog, Oophaga pumilio, shows extreme variation in color and pattern between populations that have been geographically isolated for more than 10,000 years.[22]When populations are separated by geographic distances and landscape barriers, they frequently experience restricted gene flow, which can enable phenotypic divergence between populations through selection or drift.[23] Their variety in warning coloration is used for their visibility, toxicity and resistance to predators. When divergent phenotypes are mostly restricted to separate islands, the biogeography of color polymorphism suggests a major role for neutral process. However, Summers et al. (1997)[24] provide evidences that neutral divergence alone is unlikely to have caused the variation in color patterns. As shown by Lande, rapid evolution in sexually selecting species is led by the interaction of random genetic drift with natural and sexual selection such as random genetic drift in female mating preferences.[25] Color is known to play a role in male–female signaling, mate attraction, and male–male signaling in anurans. Based on Tazzyman and Iwasa’s study that involved collections of samples from main islands in the Bocas del Toro archipelago, its results proved that female preference on male calls led to call divergence and therefore divergence was driven by sexual selection. Mate choice plays a critical role in generating and maintaining biodiversity.[26] Furthermore, spatial variation in predators or habitat features could exert divergent natural selection on coloration in response to its subjection to predator selection.[22] It is still unclear to what extent sexual selection has driven the evolution of color morphs rather than reinforcing the reproductive isolation of morphs.[27] In an aposematicorganism such as Oophaga pumilio, we cannot attribute a phylogenetic signal of selection to female mate choice alone but is quite possible that genetic drift would interact with female color preferences to trigger divergence [22]
Bastimentos color morph
Captivity :
Oophaga pumilio is a popular frog in captivity, due to its striking colors and unique life cycle. They have been imported in vast quantities to the United States and Europe since the early 1990s, when they would typically be available for around US$30 each. However, these shipments have since stopped, and O. pumilio is much less common and available in reduced diversity. In Europe, O. pumilio is much more diverse and available due to an increased frequency of smuggling and the resulting offspring of smuggled animals. Smuggling of dart frogs is less common elsewhere, but still problematic as it kills large numbers of animals and frequently degrades or destroys viable habitat.
Recently, O. pumilio has been exported from Central America again in small numbers from frog farms. Because of this, they have seen a huge increase in numbers in the dart frog community and are regularly available.
Common color morphs in captivity:
One example of a color morph is the blue jeans morph. It is most common throughout the species range, but is relatively rare in the United States pet trade. Most of these animals came from imports during the 1990s, or are their descendants.[citation needed] As of 2003, it was observed that this morph could be found throughout Costa Rica, as well as in mainland Panama.
"Blue jeans" color morph
For the external links , refrences click here to read the full wikipedia article
Video :
Oophaga pumilio "Vulture Point" calling.
Scientific. Name: Oophaga pumilio (S CHMIDT , 1857)
Etymology:
The epithet pumilio comes from the Latin word -pumilus- (= dwarf). It refers to the small size of the animals.
Synonyms:
Oophaga pumilio (G RANT , F ROST , C ALDWELL , G AGLIARDO , H ADDAD , K OK , M EANS , N OONAN, S CHARGEL & HEELER , 2006)
Oophaga pumilio (B AUER , 1994)
Dendrobates galindoi (T RAPIDO , 1953)
Dendrobates typographicus (O ERTER , 1951)
Hylaplesia typographa (B ROCCHI , 1882)
Dendrobates ignitus (C OPE , 1874)
Dendrobates typographus (K EFERSTEIN , 1867)
Hylaplesia pumilio (G ÜNTHER , 1859)
Dendrobates pumilio (O. S CHMIDT , 1857)
English name: Red-and-blue Poison Frog (CITES), Flaming Poison Frog (CITES)
German name: Panama Growler
Spanish name: Rana roja, Rana venenosa
Dutch name: Aardbeikikkertje
Classification:
Amphibia-> Anura-> Dendrobatoidea-> Dendrobatidae-> Dendrobatinae-> Oophaga -> Oophaga pumilio (S CHMIDT , 1857)
Group:
The genus Oophaga B AUER 1994 corresponds to the former Histrionicus group in the sense of M YERS (1984) or the classification of S ILVERSTONE (1975) in Pumilio and Histrionicus group.
Note on the scheme:
Recent genetic studies by H AGEMANN (2005) and R UDH (2005) and morphological data and call analyzes by K ARSCH (2004) indicate three ways within the various populations designated O. pumilio . The populations of Costa Rica, known as "blue-leger", which spread north of the Río Reventazon and the population of the Panamanian island of Isla Escudo de Veraguas, have nonemonophyleticOrigin with O. pumilio sensu strictoand therefore should be considered as two separate species. For the Northcostarican populations, the previoussynonym O. typographa (Keferstein, 1867) regain validity, the type material for thistaxonas genetically identical. Otherwise, the younger synonym Oophaga ignita (Cope, 1874) could again get Artrang, which was set up for the Nicaraguan populations. Until the final clarification we call the northern populations here as O.cf.typograph . The small red-blue populations of Isla Escudo and the mainland west of Punta Valiente are genetically much closer to O. speciosa (H AGEMANN , 2005) and also differmorphological and in theirs Anzeigerufs significantby O. pumilio (O STROWSKI , 2006). They are thus part of a new, undescribed species, which we call Oophagasp.Separate "Escudo" from O. pumilio and perform separately.






Fig .: Maximum likelihood phylogeny of the pumilio clade, based on 16 SrRNA, Cyt b and COI sequences. (CR = Costa Rica population, P = Panama population, S = southern clade, N = northern clade, E = Isla Escudo). H AGEMANN & P RÖHL(submitted), © 2007 Sabine Hagemann
Fig: Neighbor-joining tree (cytochrome b, cytochrome oxidase I and 16SrRNA). H AGEMANN , 2005.
© Sabine Hagemann & Thorsten Mahn 2006.

Threat status :
In the Red List because of its high adaptability and large-scale distribution as "low-risk" classified (IUCN, 2006).
Annex II of the WA. Annex B of EC Regulation 338/97. Notification and proof obligation according to § 6 Abs. 2 BArtSchVO.
Description :
- size:
Medium sized Oophaga species with a head-trunk length of 14 - 22 mm and a weight of 155 - 970 mg (depending on population). An examination of the body length (SVL) and the weight (BW) of 165 individuals from 11 different populations of the Bocas archipelago yielded a mean size of 18.2 mm (+/- 1.68 mm) and a mean weight of 556 mg (+/- 165 mg). There are considerable weight and size differences between the different populations (see Tab.). In O. pumilio therefore is not only apolymorphism in terms of coloring but also on body measurements. Data according to H AGEMANN (2005).
Table: Head-body length and weight (means and standard deviation) of various populations of O. pumilio . Data according to HAGEMANN (2005)



Grooming:
A clearer Sexual dimorphismis not present. Calling males show immediately after calling a dark colored area around the subgular acoustic bubble. This is more or less pronounced depending on the variant and the individual. In males who have not called for a long time (suppressed animals, animals without territory), this dark mark is often indistinct or not to see and therefore not a sure differentiator. Depending on their age and nutritional status, females usually have a slightly larger head-hull length and a somewhat fuller body (O STROWSKI , personal observation).
Fig .: Pair of Oophaga pumilio .
♂ (left), ♀ (right).
Fig .: Dorsal and ventral view of Oophaga pumilio
Variation:
The classification of the color morphs of Oophaga pumilio was based on common morphological features in coloring and drawing patterns. Differing color subpopulations within common (sympatric) distribution areas are summarized into a morpheme as a total population.
Fig .: Variability of O. pumilio "Róbalo" within a subpopulation at the site: Quebrada Cascaje.
(A) + (F) common patterns of drawing (A: Róbalo pattern with black legs, F: Uyama pattern with brown legs). (B) - (E) drawing patterns rarely found on this fund. (G) ventral of animal B. (H) ventral of animal A. © Thomas Ostrowski
Tab .: Name, size and color distribution of different populations of Oophaga pumilio
Older:
up to 15 years
Maturity:
with 12-14 months. The first clutches are often of inferior quality (fungi).
Skin poisons:
Like most of the aposematisch
colored representatives of Dendrobatinae also contains the skin of Oophaga pumilio a large number of different toxine from the class of the alkaloid
e. Many poisons isolated from poison dart frogs are among the most toxic toxins. For example, from the skin Phyllobates terribilis recovered batrachotoxin
, From the skin of O. pumilio a class of very poisonous alkaloids could be won for the first time pumiliotoxine received.
The origin of the poisons was unknown for a long time. But they are probably not produced by the frog itself, but mainly absorbed by the food, enriched and possibly slightly modified. For example, S PANDE ET AL (1999) detected two decahydroquinoline-class toxins in the body of the tropical ant Solenopsis azteca for the first time. Decahydoquinolines are one of the main venom classes in poison dart frogs and this ant species is also looted by poison dart frogs. Thus, the researchers around S PANDE then also found two stereoisomers (slightly modified form) of the ant toxins in the skin of O. pumilio, These findings support the suggestion that frogs' food is the source of the toxins.


Fig .: The tropical ant Solenopsis azteca (here at Cecropia sp. ) Contains toxins that are very similar to the skin poison of Oophaga pumilio . The red marks within the structural formulas show the deviations of the configuration from the frog poisons.
O. pumilio seems very good due to the strong skin toxin predation
to be protected. So far, there are only a few observations about natural predators of the species. Even the high age attainable in the natural biotope (P RÖHL , pers. Komm), speaks rather against a high risk of predation. Although for the sympatric occurring poison dart frog species Dendrobates auratus Observations on predation by birds and spiders are present(S UMMERS ), a predation on O. pumilio in the field has not yet been documented. The sympatric occurring, about 4 cm large predatory ant, Paraponera clavata avoids the predation on O. pumilio , although small anures belong to their natural prey spectrum(seeFig.). Feeding experiments by F RITZ ET AL . (1981) showed that whistling frogs ( Eleutherodactylus ssp .) Belong to the prey spectrum of the ant, are immediately attacked and completely consumed. In contrast, the approximately the same size wasO. pumilio in the experiment either not attacked or immediately released. S ZELISTKOWSKI (1985) reported almost identical observations on the hunting behavior of the spider Cupiennius coccineus . This also captures both frog species, the Eleutherodactylus eats but quickly releases O. pumilio from the clutches. The poison of the species thus seems to have a deterrent effect on large predatory arthropods.




Pic .: The tropical ant Paraponera clavata is a danger with 4 cm length even for smaller Anuren. Here it consumes a Eleutherodactylus ridens . She avoids contact
with O. pumilio . © 2006 Tobias Eisenberg www.t-eisenberg.de
Fig .: Parasitic attack in Oophaga pumilio . Nematodes in muscle tissue.
© 2005 Sabine Hagemann
Fig .: Predatory spiders of the genus Cupiennius also capture smaller vertebrates. But as predators to O. pumilio they are probably out of the question. © Thomas Ostrowski 2007
Fig .: Larva of a fly in the muscle tissue of Oophaga pumilio © 2005 Sabine Hagemann
Clutch and larvae :




Fig .: Scrim of Oophaga pumilio
© 2005 Sabine Hagemann
Nutrition:
Like all members of the genus Oophaga , the larvae of Oophaga pumilio only take on the nourishing eggs produced by the female as food. In the phytotelmata
of the natural breeding plants contain a multiplicity of organisms, which could serve as source of food (mosquito and other insect larvae, algae, unicellular organisms, detritus
etc.). Nevertheless, larvae of O. pumilio starve to death when the caring female fails, for example through death. The larvae are thus highly dependent on the nutrient supply by their mother (obligatory oophag). The specialization goes even further. During the supply of larvae, the female produces physiologically special eggs that are only used for larval feeding (Abortiveier). They have a different composition and no particularly pronounced jelly. They can not be fertilized and do not develop into larvae. During the brood care period, females of O. pumilio do not lay any fertile clutches. Before W EYGOLD (1980) was able to document the complex brood care of the species for the first time, several attempts were made to artificially raise larvae of O. pumilio . The food problem causes great difficulties. B ECHTER (1978) was finally able to find larvae of O. pumiliowith egg yolks as substitute food. The yolk is added dropwise and eaten by the larvae. However, since egg yolk spoils the water quality quickly, a water change must be made shortly after feeding. In addition to the elaborate procedure but it also comes to developmental problems. These manifest themselves in prolonged developmental periods and / or malformations. B EUTELSCHIESS (1983) was able to test another substitute feed with O. speciosa with a mixture of marmalade quark, egg yolk, soy flakes and osspulvite (vitamin / mineral mixture). However, the developmental disorders described above also occurred. To date, only eggs from other Dendrobatidae come into question as suitable replacement feed. The author could larvae of the Rambala variant ofO. pumilio with the eggs of Ranitomeya cf. ventrimaculata successfully raise. Neither developmental disturbances occurred nor could time delays in the ontogenesis be determined. The little frogs ended theirs metamorphosis After about 40 days, it took about as long as larvae reared by the mother (O STROWSKI , personal observation). However, the eggs must first be freed from the gelatinous hull, as the larvae can not recognize the Fremdgelege otherwise as food. For this purpose, the jelly must be cut and the urchins are sucked from the jelly (pipette). Every 2-3 days a larva received such a replacement. This type of breeding is quite elaborate and requires another Pfeilgiftfroschart in attitude that regularly produces eggs. For example, Phyllobates lugubris, Epipedobates anthonyi or R. variabilis are possible . However, the simplest and most effective method is the natural breeding by the mother.
Behavior (ethology) :
- General behavior:
Oophaga pumilio is a diurnal inhabitant of the foliage layer. The frogs live mainly ground-oriented. But you can climb well and use, in the absence of near-ground breeding structures, including the epiphyte enflora on trees at higher altitudes, around larvae in small phytotelmata raise. Foraging and courtship, however, usually take place on the ground. The species O. pumilio is not shy and active almost all day long. Only on very hot days can no calls be heard in the biotope at lunchtime. The species has a very complex territorial, courtship and brood care behavior and has therefore been one of the most popular behavioral biological research objects among herpetologists for decades. Countless puplications have been published so far, making O. pumilio the most studied tropical anuran species ever. Several puplications refer to populations from Costa Rica, possibly as the O. typographa of O. pumiliohave to be split off (see systematics). However, the different populations differ only slightly in their behavior. significant ethologically
Distinguishing features are found mainly in the physical parameters of their Anzeigeruf
e. Therefore, assigning the puplications to different populations and thus distinguishing the behavior does not seem to make sense. The statements made here apply both to the populations of Costa Rica ( O. pumilio sl, possibly O. typographa ) as well as on the Panamanian populations ( O. pumilio ss) too.
- Territorial:
O. pumilio is an extraordinarily well- behaved frog (B UNNEL , 1973). Certain males could over several years again and again in their ancestral territoryor even be observed at their preferred call location (P RÖHL , pers. Komm.). Studies by D ONNELLY (1989) showed that O. pumilio Aktionsräume (Home-range) occupied from 2 to 15 square meters. The actual territory (defended area) is smaller. Males occupy smaller spaces of action (2.26-15.07 square meters) than female animals (5.72-15.11 square meters). This gender inequality could have several causes. For example, females may have an increased need for feed as they are responsible for the more energy-consuming part of the reproduction (larval feeding with Abortiveiern), because they require different habitat structures than males for breeding success (eg phytotelmata) or because female intervention rooms overlap with multiple male action rooms to allow better mate choice. Such overlapping zones also appear much less frequently between the action spaces of males than between the action spaces of several females or between females and males. The core zones of the male action spaces, ie the actual territories, are usually 2-3 m, rarely up to 6 m, apart (B UNNEL , 1973). This gender specificity indicates a less pronounced intrasexual aggression in female D. pumilioout.


Male wrestling
Female eats foreign nest.
While aggressive behaviors between males can be observed in the field, they have not been described in females. In the terrarium and under laboratory conditions, however, aggressive behaviors could also be observed among female animals (W EYGOLD, 1984). This rarely leads to wrestling matches and more often to eating foreign material from other females (see photo). At the meeting of male animals at the territorial boundaries, it first comes to Rufduellen in which the frequency and call rate of Anzeigerufe be modulated to intimidate the opponent and keep from entering the own territory. In case of ineffectiveness of these reputation duels and the penetration of an equally strong enemy into the foreign territory, it then comes very quickly to wrestling matches among the males (B UNNELL, 1973). The opponents try to throw each other on the back and then to press on the ground (s.Foto). The subject frog acts then completely passive and flees as soon as the winner leaves him again. He is pursued by the dominant male under constant call for a while (usually over the territorial boundaries). If the inferior opponent of the territory owner, so the action space limits can now move and pull in the worst case, the loss of the area and takeover by the enemy. However, the Revierinhabr usually has a home advantage and wins such wrestling matches.
Amazing is also the ability to orient when retrieving their own territory. After being removed from their ancestral territory and released from the area, most of O. pumilio find their own territory, even many meters away, in a targeted manner. These are often unknown areas outside of their ownhome range it traverses it and several foreign territories of O. pumilio (O STROVSKI , pers. Observ .). An amazing feat for a frog of only about 2 cm in length. Alone visual orientation to terrain landmarks can play only a minor role in such a small frog and a highly structured, unclear biotope. It can only be significant within the known (relatively small) action space. Attempts by F ORESTER & W ISNIESKI (1991) indicate that the sense of smell plays an important role in the recognition of one's own territory. In the experiment, the frogs preferred two offered odors significant the smell of their ancestral territory, the inhabited terrarium. Above all, volatile chemical substances that are released from the plants (eg bromeliads) of the inhabited habitat, appear in the olfactory orientation play an important role. These smells paint a characteristic picture of the environment and seem to be used by the frog as orientation in the re-finding of the ancestral territory, in addition to the spatial orientation. In the experiment, the frogs were able to recognize their territory solely because of the "native" smell.
- courtship behavior:






Couple at the Balz © 2007 Gert Benaets
- Brood care behavior:
Calling male recruits a female ready to mate © 2013 Leonard Georg
At regular intervals, the female returns to larvae-occupied Phytotelme. © 2007 Thomas Ostrowski
A couple Oophaga pumilio shortly before oviposition
© 2008 Gert Benaets
Calling male recruits a female ready to mate © 2013 Leonard Georg
Females of O. pumilio transporting a larva.
© 2007 Thomas Ostrowski


A few days old cub explores the biotope
© 2013 Leonard Georg
Freshly metamorphosed frogs of Oophaga pumilio usually remain in the phytotelm for several days before they leave. © 2007 Thomas Ostrowski
- Utterance (vocalization): Click here
Habitat :
Type find location of the first description
"the border area between New Granada [Colombia with province Panama] and Costa Rica, the path between Bocca del toro and the Vulcan Chiriqui [Panama] ... between 5000' and 7000' height" [Polish feet = 1150-1160 m ]. (S CHMIDT , 1858)

Distribution area Oophaga pumilio
© 2008
Thorsten Mahn
Distribution:
As a lowland species at altitudes 0-500 m [800 m to S AVAGE , 2001] along the Caribbean coast of the Isthmus from central Costa Rica (Río Reventazon) to western Panama (Archipelago de Bocas del Toro).
The altitude of 800 m in the literature, the author seems too high and could be due to a Fehlmessung (or estimate) of the Erstbeschreibers. S CHMIDT (1855) collected the species in the province of Bocas del Toro and from there crossed the Cordillera de Talamaca towards Bouquete. He also collected the species O. speciosa,
For both species he gives heights of 800 m for the localities. This was more likely to be the pass of his route. The author was unable to detect populations in Panama or Costa Rica at over 500 m (O STROWSKI , personal observation).
Biotope:
Oophaga pumilio originally inhabited Caribbean lowland rainforests at altitudes of up to 500 m. In semi-natural forests, the species lives in the deciduous and herbaceous layer and rarely climbs above 1 m in height. It is not an arboreal (tree-inhabiting) species. O. pumiliois extremely adaptable. In the natural habitat, the species inhabits tropical lowland rainforest, evergreen semi-moist forest, found in mango forests or as on the islands of the Bocas Archipelago along the edge of the forest, near the beach in the influence of salty spray. Due to its natural adaptability, the species also copes well with anthropogenic areas and even cultural landscapes. We were able to detect the species at ditches, in gardens, parks, on cattle pastures, in extensively cultivated banana and pineapple plantations, in overgrown cemeteries, and even in small rubbish dumps close to settlements.












On the Isla Bastimentos O. pumilio even populates the alluvial garbage directly at the rinsing seam. Prov. Bocas del Toro, Panama © 2006 Holger Birkhahn
On the island Bastimentos O. pumilio can be found along the edge of the forest near the beach. Prov. Bocas del Toro, Panama © 2006 Thomas Ostrowski
Dry palm fronds on the forest floor on the Isla Solarte. Biotope of the "Solarte" variant.
Panama, province Bocas del Toro. © 2004 F. Steinmann
Undergrowth from Heliconia sp. and Dieffenbachia sp. in the biotope of the "Bastimentos" variant.
Panama, province Bocas del Toro. © 2005 T. Ostrowski
An extensively managed banana plantation.
Biotope of the "Guarumo" variant.
Panama, province Bocas del Toro. © 2005 T.Ostrowski
Along the banks of the Rio Cataratas there is a large population of the Bri Bri variety. Prov. Limón, Costa Rica. © 2006 Holger Birkhahn
This meadow at the edge of a cocoa plantation is inhabited by the blue "Tierra Oscura" variety.
Panama, province Bocas del Toro. © 2005 T. Ostrowski
The old cemetery of Punta Hospital. Numerous populated with animals of the "Solarte" variety.
Panama, province Bocas del Toro. © 2005 S. Hagemann
Dense stocks of Dieffenbachia sp. on Isla Bastimentos. Biotope of the "Bahia Honda" variant.
Panama, province Bocas del Toro. © 2005 T. Ostrowski

Biotope of the "Solarte" variant
Panama, province Bocas del Toro. © 2006 H. Birkhahn
Biotope Colón
In Baumbusstauden is the habitat of Oophaga pumilio on Isla Colón
Along the banks of small Quebradas in the protected area Hitoy Cerere there are dense populations of breeding plants for O. pumilio . Prov. Limón, Costa Rica.
Another advantage of clearing areas as a preferred habitat could also be the increased feed density. Scraping organic material on clearing areas or in rubbish sites could attract significantly more potential feed insects. Thus, even at refuse sites often larger population densities of O. pumilio and Dendrobates auratus can be detected. However, food availability appears to have a much smaller impact on the population structure. Increased densities of D. auratus even on very old refuse sites that were no longer used were mostly a result of increased artificial accumulation of water and not increased feed density (O STROWSKI , personal observation).
Climate:
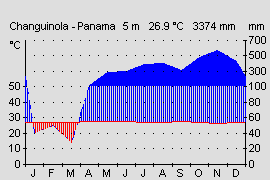
The southern end of the isthmus(Costa Rica and Panama) is climatically separated by the Cordilleras into a humid Caribbean and a drier Pacific side. The Caribbean side of Panama is characterized by a fairly even tropical climate. In the course of the year, the temperature fluctuations are very low and average about 1-2 °. Even in the course of the day, higher variations between day and night can rarely be detected in the Caribbean lowlands (W ALTER & B RECKLE, 1999). However, on clear nights, after heavy rains or Atlantic cold air currents due to hurricanes in the Caribbean Sea can lead to short-term temperature drops. In the autumn of 2004 in Changuinola, Panama at sea level, a cold snap with temperatures of only 15 ° C. Also in Costa Rica, the author was after heavy rainfall in the lowlands temperatures of 16 ° C measure (O STROWSKI , pers. Observed). However, these unusual low temperatures last only a few hours (in exceptional cases a few days) and are usually only locally limited. On the protected forest floor, however, temperatures can be measured almost year-round, which correspond approximately to the annual average both during the day and at night (W ALTER & BRECKLE , 2000). In the province of Bocas del Toro this is about 26 ° C (see Fig.). The rainfall is also relatively constant and high year round. During the northern winter, the Northeast Passage's air currents bring humid air masses, while in the northern summer, low pressure areas over the Caribbean cause precipitation. The precipitation varies between 2000 and 4000 mm per year, depending on the shadow from the mountain ranges (W ALTER & B RECKLE , 1999). Only in the months of January to March can there be short dry periods of less than 100 mm per month (see Fig.). Overall, the rainfall over the year but relatively constant, so that the frogs on the Caribbean side can reproduce throughout the year.
Especially important is the presence of breeding possibilities. The nature of the biotope is determined by the number of possible larval distribution sites (phytotelmata) directly the population structure. P RÖHL (2002) was able to prove in her studies that the population density in the primary forest is relatively low and that a close to balanced operational gender ratio (=OSR) of 1: 1 prevails. In the more open secondary forest an OSR shifted to the female side could be detected and the total population density was significantly higher. These results were always correlated with the increased presence of breeding possibilities (in this case Heliconia and Dieffenbachien). O. pumilio is a ground-based frog and prefers to use breeding plants from the Helikonia (Heliconiaceae) and Araceae family. Although these plants are relatively undemanding and little hungry for light, they can only thrive in the primary forest at light-exposed areas, such as in the area of tree cover, wind break or on river banks. Thus, the availability of suitable districts for courtship and brood care in primary forest is limited to such natural islands of light.epiphytes The treetops can also be used only rarely, since in the primary forest of thelowlands only in the light flooded crowns freestanding About stand, larger amounts of epiphytes with Phytotelmata (Bromeliaceae) occur.
Areas with more light (secondary forest, plantations, clearing areas or roadsides) allow a better growth of the ground vegetation and the epiphyte flora also at lower root areas. This increases the number of potential breeding plants and directly increases the population density of O. pumilio. The author was able to visit a clearing area in Costa Rica and observe the course of the population structure over two years. The distances between individual territories of O. pumilio sl ( O. cf. typographa) were in the primary forest at over 10 m. The animals were usually associated in pairs. On the adjacent clearing area, the distance between the animals was often less than 2 m and it could be detected on a male about three females. Due to the increasing presence of bromeliads on the ground (on cleared trees), the population grew strongly within one year. In addition, animals migrated from the adjacent forest. Alongside roads, dense heliconia stands can be found in many places, often serving as a biotope for a large O. pumiliopopulation.

Attitude in the terrarium :
Terrarium / Facility:
Rainforest terrarium from 50x50x50cm
automatic irrigation and fog plant recommended Bodenbewohner
, climbs often and much
Temperatures:
24-27 ° C, drop by 3-4 ° C at night
Annual variation: keep drier in winter Yearly variation in
temperature minimum (1-2 ° C)
Humidity:
70-80%, at lunchtime up to 70%, in the morning and in the evening 100% (fog)
Annual variation: Rainy season with high humidity and rain between May and September
Nutrition:
Usual small to medium food animals like
- springtails
.- Small fruit fly
.- Big fruit fly
.- Micro grilling
- crickets,
- Small waxmead
- White Assel
- barklice
- (Micro beetle), and finely sifted
- meadow plankton
, For juveniles, springtails are essential in the first four to six months. For Adult Feeding animals such as crickets and fruit flies should be dusted regularly 1-2 times a week with a good vitamin preparation (eg Amivit A according to the original recipe of B IRKHAHN, 1991). Feed animals for young animals should be pollinated daily for the first 4 weeks. Store opened vitamin supplements in the fridge. Fruit flies can be well fed before feeding with liquid vitamin preparations (eg Sanostol, Multibionta) and so be nutritionally and nutritionally valuable. Pollinated food animals should be offered in the terrarium on exchangeable trays. Residual vitamin powder residues can not form a bacterial focus on the terrarium floor. Fruit pieces designed in small bowls in the terrarium (eg banana slices) are good places for fruit flies and are soon accepted by the frogs as feeding places. For a sufficient vitamin supply of the feed animals by these lures the dwell time of the feed animals should be however too small, so that should be additionally vitaminized. Offered food trays should be cleaned every 2-3 days for hygienic reasons. Spring tails can be well focused on laid out Xaxim pieces by using them with small! Quantities of dry yeast sprinkled. Here, too, the frogs quickly learn the meaning of the feeding place.
trim:
Good results were achieved with the combinations 1,1 and 1,2. In several males it comes to oppression rituals. In large pools also group housing possible. Too many old animals complicate the young rearing.
Tips for breeding:
Oviposition occurs on smooth leaves rarely in caves.
Lays 5-10 eggs
Development time Eggs: 10-14 days
Active transport of the burbot by the female into very small accumulations of water
Development of the tadpoles: 80-90 days water temperature at 25 ° C, night setback recommended
The burbots are supplied by the female with special feeders, artificial rearing with substitute eggs of dendrobatids of the same species possible, but laborious. Eggs of the species D. auratus, D. tinctorius, P. vittatus, P. bicolor, P. lugubris and E. tricolor are suitable for feeding. Since O. pumillio larvae can not eat the jellies of foreign eggs, they must be removed before feeding without destroying the fruit. With a small curved forceps and on the dry it is easiest. The development time of the tadpole is longer in the rearing with foreign eggs and the failure rate is higher! Exception: in R. ventrimaculata eggs the rearing takes only 10 weeks, feeding up to 2 times a week.
Therefore, the breeding of the burbot and juvenile is best done with the parents in the breeding tank. Early kittens (not before 6 months) often stop feeding and die.
For more information about resources for the above article .. click here
Madagascar Dart frogs
Aromobatidae :
-
Allobatinae
-
Anomaloglossinae
-
Aromobatinae
South America Dart Frogs - Species
Dendrobatidae :
-
Colostethinae
-
Dendrobatinae
-
Hyloxalinae
Oophaga Genus :
Species :
1- Oophaga pumilio - Schmidt, 1857 - Strawberry poison-dart frog
Morphs Part One .. Two .. Three....
2- Oophaga granulifera - Taylor, 1958 - The granular poison frog
Part 1 .. Part 2 .. Part 3 .. ..
3- Oophaga histrionica - Berthold, 1845 - The harlequin poison frog
Part 1 .. Part 2 .. Part 3 .. ..
4- Oophaga lehmanni - Myers and Daly, 1976 - Lehmann's poison frog
5- Oophaga sylvatica - Funkhouser, 1956 :
Part 1 .. Part 2 .. Part 3 .. Part 4 ..
Oophaga Genus :
Species :
1- Oophaga pumilio - Schmidt, 1857 - Strawberry poison-dart frog
Morphs Part One .. Two .. Three....
2- Oophaga granulifera - Taylor, 1958 - The granular poison frog
Part 1 .. Part 2 .. Part 3 .. ..
3- Oophaga histrionica - Berthold, 1845 - The harlequin poison frog
Part 1 .. Part 2 .. Part 3 .. ..
4- Oophaga lehmanni - Myers and Daly, 1976 - Lehmann's poison frog
5- Oophaga sylvatica - Funkhouser, 1956 :
Part 1 .. Part 2 .. Part 3 .. Part 4 ..